
Valorisation of Bovine Sweet Whey and Sunflower Press Cake Blend through Controlled Fermentation as Platform for Innovative Food Materials

 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Identification of Micro-Organisms
2.2. Strain Discrimination by Molecular Fingerprinting
2.3. Determination of Phytase Activity and Carbohydrates Assimilation Profile of the Strains
2.4. Growth Test in Bovine Sweet Whey
2.5. Fermentation Trials in Microcosms
2.6. Determination of Sugars and Organic Acids by HPLC Method
2.7. Statistical Analysis
3. Results
3.1. Micro-Organism Identification and Strain Typing
3.2. Phenotypic Characterization of Strains
3.3. Growth Test in Bovine Sweet Whey
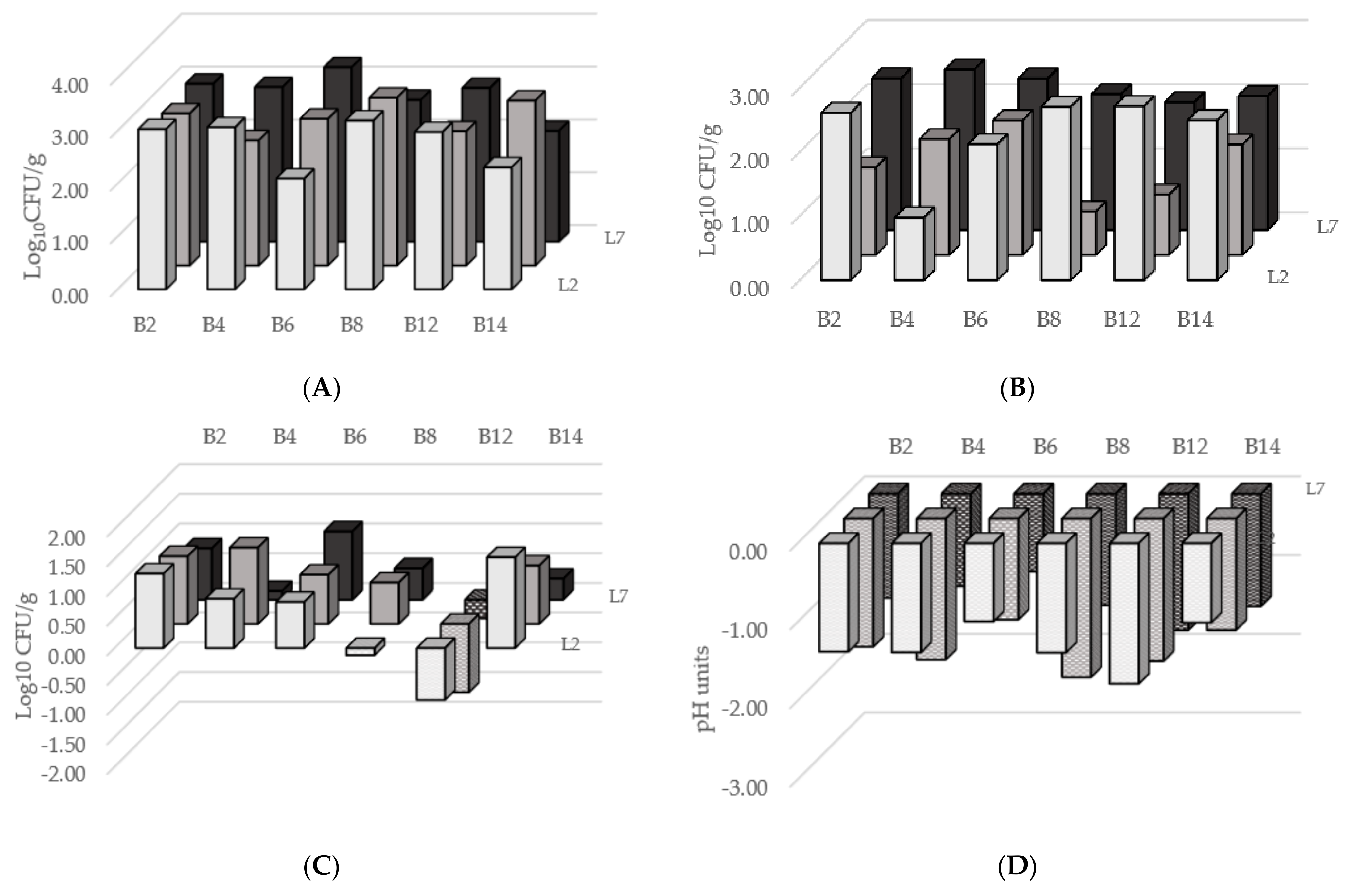
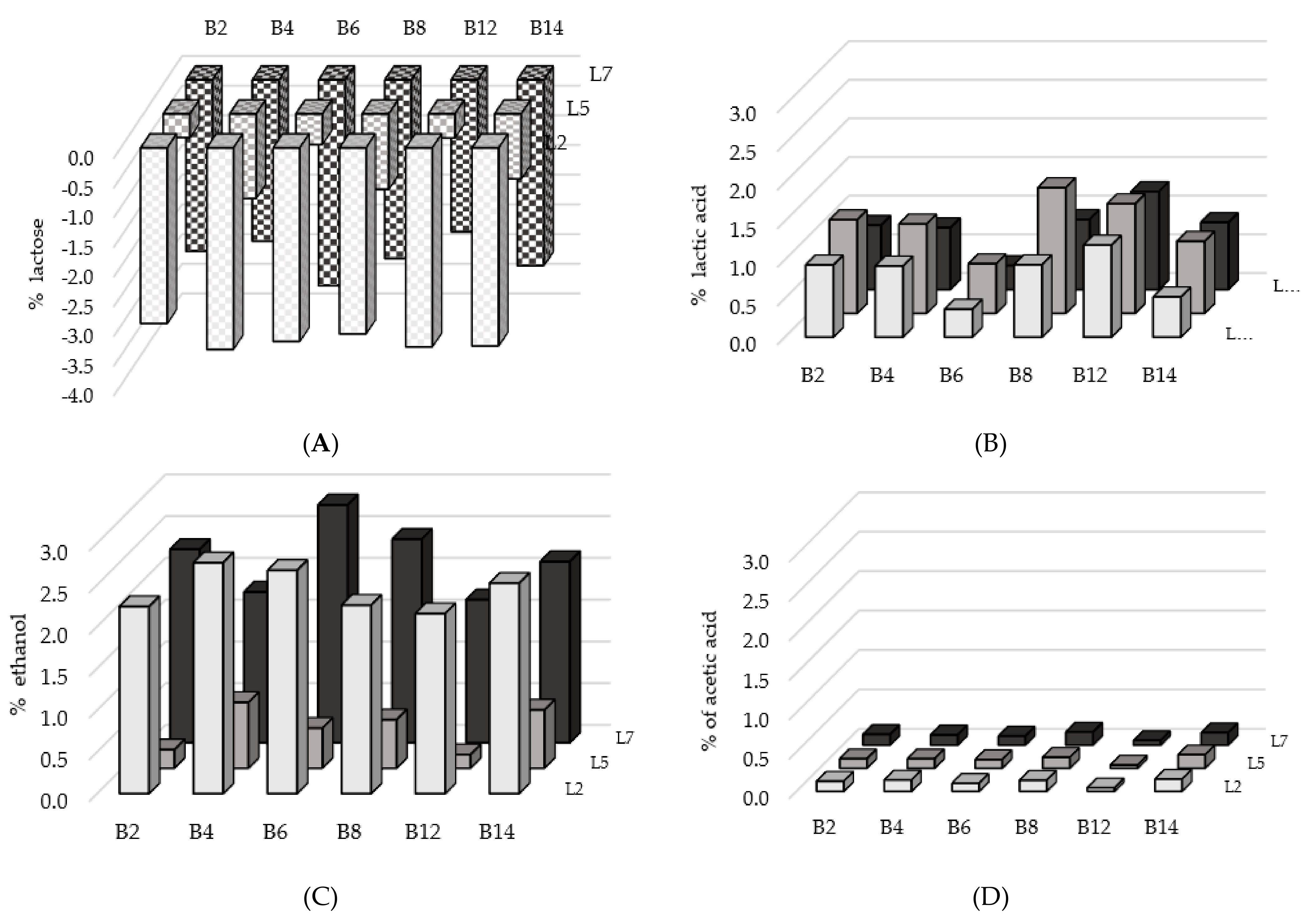
3.4. Fermentation Trials in Microcosms
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- European Commission. Communication from the Commission to the European Parliament, the European Council, the Council, the European Economic and Social Committee and the Committee of the Regions; The European Green Deal. Bruxelles 11.12.2019 COM(2019) 640 Final; European Commission: Brussels, Belgium, 2019. [Google Scholar]
- Winkler, H. Closed-loop production systems—A sustainable supply chain. CIRP J. Manuf. Sci. Technol. 2011, 4, 243–246. [Google Scholar] [CrossRef]
- Directive 2008/98/EC of the European Parliament and of the Council of 19 November 2008 on waste and repealing certain directives. Off. J. Eur. Union 2008, L 312, 3–30.
- Moates, G.; Sweet, N.; Bygrave, K.; Waldron, K. Top 20 Food Waste Streams. REFRESH Project Deliverable 6.9 WRAP/IFR. 2016. Available online: https://eu-refresh.org/top-20-food-waste-streams.html (accessed on 12 April 2022).
- Rao, M.; Bast, A.; de Boer, A. Valorized food processing by-products in the EU: Finding the balance between safety, nutrition, and sustainability. Sustainability 2021, 13, 4428. [Google Scholar] [CrossRef]
- Eurostat. Agriculture, Forestry and Fishery Statistics; Publications Office of the European Union: Luxembourg, 2020. [Google Scholar] [CrossRef]
- Paraman, I.; Sharif, M.K.; Supriyadi, S.; Rizvi, S.S.H. Agro-food industry by-products in to value-added extruded foods. Food Bioprod. Proc. 2015, 96, 78–85. [Google Scholar] [CrossRef] [Green Version]
- Lappa, I.K.; Papadaki, A.; Kachrimanidou, V.; Terpou, A.; Koulougliotis, D.; Eriotou, E.; Kopsahelis, N. Cheese whey processing: Integrated biorefinery concepts and emerging food applications. Foods 2019, 8, 347. [Google Scholar] [CrossRef] [Green Version]
- Petraru, A.; Amariei, S. Oil press-cakes and meals valorization through circular economy approaches: A review. Appl. Sci. 2020, 10, 7432. [Google Scholar] [CrossRef]
- Srilatha, K.; Krishnakumari, K. Proximate composition and protein quality evaluation of recipes containing sunflower cake. Plant Foods Hum. Nutr. 2003, 58, 1–11. [Google Scholar] [CrossRef]
- Weisz, G.M.; Kammerer, D.R.; Carle, R. Identification and quantification of phenolic compounds from sunflower (Helianthus annuus L.) kernels and shells by HPLC-DAD/ESI-MSn. Food Chem. 2009, 115, 758–765. [Google Scholar] [CrossRef]
- Tangyu, M.; Muller, J.; Bolten, C.J.; Wittmann, C. Fermentation of plant-based milk alternatives for improved flavour and nutritional value. Appl. Microbiol. Biotechnol. 2019, 103, 9263–9275. [Google Scholar] [CrossRef] [Green Version]
- Akal, C. Chapter 28—Benefits of whey proteins on human health. In Dairy in Human Health and Disease across the Lifespan; Watson, R.R., Collier, R.J., Preedy, V.R., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 363–372. [Google Scholar] [CrossRef]
- Querol, A.; Barrio, E.; Huerta, T.; Ramon, D. Molecular monitoring of wine fermentations conducted by Active Dry Yeast strains. Appl. Environ. Microbiol. 1992, 58, 2948–2953. [Google Scholar] [CrossRef] [Green Version]
- Green, M.R.; Sambrook, J. Molecular Cloning: A Laboratory Manual, 4th ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2012. [Google Scholar]
- Edwards, U.; Rogall, T.; Blöcker, H.; Emde, M.; Böttger, E.C. Isolation and direct complete nucleotide determination of entire genes. Characterization of a gene coding for 16S ribosomal RNA. Nucleic Acids Res. 1989, 17, 7843–7853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vigentini, I.; Praz, A.; Domeneghetti, D.; Zenato, S.; Picozzi, C.; Barmaz, A.; Foschino, R. Characterization of malolactic bacteria isolated from Aosta Valley wines and evidence of psychrotrophy in some strains. J. Appl. Microbiol. 2016, 120, 934–945. [Google Scholar] [CrossRef] [PubMed]
- Kurtzman, C.P.; Robnett, C.J. Identification and phylogeny of ascomycetous yeasts from analysis of nuclear large subunit (26S) ribosomal DNA partial sequences. Antonie van Leeuwenhoek 1998, 73, 331–371. [Google Scholar] [CrossRef] [PubMed]
- Rossetti, L.; Giraffa, G. Rapid identification of dairy lactic acid bacteria by M13-generated, RAPD-PCR fingerprint databases. J. Microbiol. Methods 2005, 63, 135–144. [Google Scholar] [CrossRef]
- Freitas, D.B.; Reis, M.P.; Lima-Bittencourt, C.I.; Costa, P.S.; Assis, P.S.; Chartone-Souza, E.; Nascimento, A.M. Genotypic and phenotypic diversity of Bacillus spp. isolated from steel plant waste. BMC Res. Notes 2008, 1, 92. [Google Scholar] [CrossRef] [Green Version]
- Cordero-Bueso, G.; Mangieri, N.; Maghradze, D.; Foschino, R.; Valdetara, F.; Cantoral, J.M.; Vigentini, I. Wild grape-associated yeasts as promising biocontrol agents against Vitis vinifera fungal pathogens. Front. Microbiol. 2017, 8, 2025. [Google Scholar] [CrossRef] [Green Version]
- Raghavendra, P.; Halami, P.M. Screening, selection and characterization of phytic acid degrading lactic acid bacteria from chicken intestine. Int. J. Food Microbiol. 2009, 133, 129–134. [Google Scholar] [CrossRef]
- Nielsen, M.M.; Damstrup, M.L.; Hansen, Å. An optimised micro-titer plate method for characterisation of endogenous rye phytase under industrial rye bread making conditions. Eur. Food Res. Technol. 2008, 227, 1009–1015. [Google Scholar] [CrossRef]
- Altilia, S.; Foschino, R.; Grassi, S.; Antoniani, D.; Dal Bello, F.; Vigentini, I. Investigating the growth kinetics in sourdough microbial associations. Food Microbiol. 2021, 99, 103837. [Google Scholar] [CrossRef]
- El Hag, M.E.; El Tinay, A.H.; Yousif, N.E. Effect of fermentation and dehulling on starch, total polyphenols, phytic acid content and in vitro protein digestibility of pearl millet. Food Chem. 2002, 77, 193–196. [Google Scholar] [CrossRef]
- Gupta, R.K.; Gangoliya, S.S.; Singh, N.K. Reduction of phytic acid and enhancement of bioavailable micronutrients in food grains. J. Food Sci. Technol. 2015, 52, 676–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Özer, B.; Kirmaci, H.A. Fermented Milks, products of Eastern Europe and Asia. In Encyclopedia of Food Microbiology, 2nd ed.; Batt, C.A., Tortorello, M.L., Eds.; Academic Press: Cambridge, MA, USA, 2014; pp. 900–907. [Google Scholar] [CrossRef]
- Van Vyk, J. Kefir: The Champagne of fermented beverages. In Fermented Beverages; Grumezescu, A.M., Holban, A.M., Eds.; Woodhead Publishing: Cambridge, UK, 2019. [Google Scholar] [CrossRef]
- Nguela, M.J.; Vernhet, A.; Julien-Ortiz, A.; Sieczkowski, N.; Mouret, J.-R. Effect of grape must polyphenols on yeast metabolism during alcoholic fermentation. Food Res. Int. 2019, 121, 161–175. [Google Scholar] [CrossRef] [PubMed]
- Parkar, S.G.; Stevenson, D.E.; Skinner, M.A. The potential influence of fruit polyphenols on colonic microflora and human gut health. Int. J. Food Microbiol. 2008, 124, 295–298. [Google Scholar] [CrossRef] [PubMed]
- Fritsch, C.; Heinrich, V.; Vogel, R.F.; Toelstede, S. Phenolic acid degradation potential and growth behavior of lactic acid bacteria in sunflower substrates. Food Microbiol. 2016, 57, 178–186. [Google Scholar] [CrossRef]
- Motarjemi, Y. Impact of small scale fermentation technology on food safety in developing countries. Int. J. Food Microbiol. 2002, 75, 213–229. [Google Scholar] [CrossRef]
- Petraru, A.; Ursachi, F.; Amariei, S. Nutritional characteristics assessment of sunflower seeds, oil and cake. Perspective of using sunflower oilcakes as a functional ingredient. Plants 2021, 10, 2487. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Code | Origin | Collection Number | Species | Identification (%) | U 1 |
|---|---|---|---|---|---|
| B1 | Homemade sugary kefir | LAC CAS 34 | Lacticaseibacillus paracasei | 99.6 | 17.1 |
| B2 | Homemade sugary kefir | LAC YAM 2 | Lacticaseibacillus paracasei | 99.8 | 20.6 |
| B3 | Homemade sugary kefir | DTB-1B | Lacticaseibacillus paracasei | 99.9 | 14.8 |
| B4 | Homemade sugary kefir | LAC CAS 32 | Lactobacillus casei | 99.6 | 18.0 |
| B5 | Homemade milk kefir | KMSG-22B | Lactobacillus kefiranofaciens | 97.1 | 19.7 |
| B6 | Homemade milk kefir | KMSG-1B | Lentilactobacillus parakefiri | 100.0 | 16.6 |
| B7 | Homemade milk kefir | KMSS-1B | Lentilactobacillus kefiri | 100.0 | 12.8 |
| B8 | Homemade milk kefir | DGL-1B | Lacticaseibacillus paracasei | 100.0 | 23.4 |
| B9 | Homemade milk kefir | DGL-2B | Lacticaseibacillus paracasei | 100.0 | 18.4 |
| B10 | Commercial milk kefir | 2KB-1B | Lactococcus lactis | 99.7 | 20.8 |
| B11 | Commercial milk kefir | 3KS-1B | Lactococcus lactis | 100 | 12.8 |
| B12 | Commercial milk kefir | 4BK-12B | Lactococcus lactis | 100.0 | 10.2 |
| B13 | Homemade sugary kefir | LAC FRU 1 | Leuconostoc citreum | 99.5 | 20.3 |
| B14 | Commercial milk kefir | 1KP-1B | Leuconostoc pseudomesenteroides | 99.4 | 6.6 |
| L1 | Commercial milk kefir | 2KB-12Y | Debaryomyces hansenii | 99.6 | 7.4 |
| L2 | Household milk kefir | AVY | Kluyveromyces lactis | 98.3 | 7.3 |
| L3 | Household milk kefir | CAND 22 | Kluyveromyces marxianus | 100.0 | 7.4 |
| L4 | CBS collection (milk kefir) | CBS 834 | Kluyveromyces marxianus | 100.0 | 7.5 |
| L5 | Commercial milk kefir | 1KP-1Y | Torulaspora delbruecki | 99.6 | 6.6 |
| L6 | Commercial milk kefir | 4BK-1Y | Pichia fermentans | 100.0 | 7.8 |
| L7 | CBS collection (yoghurt) | CBS 397 | Kluyveromyces marxianus | 99.2 | 7.2 |
| L8 | Commercial milk kefir | 3KS-1Y | Pichia fermentans | 99.6 | 7.8 |
| L9 | CBS collection (creamery) | CBS 2359 | Kluyveromyces lactis | 99.8 | 7.2 |
| L10 | Household sugary kefir | DTB-1Y | Pichia kluyveri | 99.6 | 7.4 |
| L11 | Household milk kefir | DGL-1Y | Saccharomyces cerevisiae | 99.0 | 7.3 |
| L1 | L2 | L3 | L4 | L5 | L6 | L7 | L8 | L9 | L10 | L11 | Average | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| B1 | 0.24 ± 0.10 | 0.29 ± 0.10 | 0.76 ± 0.53 | 0.35 ± 0.06 | 0.33 ± 0.14 | 0.28 ± 0.24 | 0.52 ± 0.28 | 0.16 ± 0.12 | 0.53 ± 0.48 | 0.36 ± 0.24 | 0.35 ± 0.41 | 0.36 abc ± 0.30 |
| B2 | 0.21 ± 0.04 | 0.36 ± 0.18 | 0.68 ± 0.29 | 0.60 ± 0.29 | 0.27 ± 0.15 | 0.62 ± 0.41 | 0.44 ± 0.04 | 0.20 ± 0.14 | 0.64 ± 0.63 | 0.03 ± 0.01 | 0.38 ± 0.13 | 0.39 abc ± 0.32 |
| B3 | 0.22 ± 0.13 | 0.33 ± 0.05 | 0.36 ± 0.09 | 0.54 ± 0.30 | 0.33 ± 0.09 | 0.08 ± 0.08 | 0.40 ± 0.03 | 0.16 ± 0.07 | 0.53 ± 0.27 | 0.25 ± 0.21 | 0.61 ± 0.01 | 0.33 ab ± 0.21 |
| B4 | 0.14 ± 0.06 | 0.32 ± 0.20 | 0.33 ± 0.07 | 0.43 ± 0.11 | 0.29 ± 0.05 | 0.09 ± 0.03 | 0.53 ± 0.11 | 0.21 ± 0.13 | 0.40 ± 0.21 | 0.19 ± 0.16 | 0.67 ± 0.29 | 0.34 ab ± 0.21 |
| B5 | 0.19 ± 0.13 | 0.30 ± 0.06 | 0.39 ± 0.09 | 0.33 ± 0.09 | 0.38 ± 0.12 | 0.42 ± 0.35 | 0.33 ± 0.06 | 0.51 ± 0.36 | 0.32 ± 0.10 | 0.11 ± 0.11 | 0.57 ± 0.47 | 0.34 ab ± 0.21 |
| B6 | 0.12 ± 0.05 | 0.42 ± 0.15 | 0.48 ± 0.21 | 0.33 ± 0.09 | 0.40 ± 0.18 | 0.33 ± 0.06 | 0.68 ± 0.55 | 0.27 ± 0.19 | 0.98 ± 0.22 | 0.14 ± 0.06 | 0.31 ± 0.06 | 0.40 abc ± 0.30 |
| B7 | 0.10 ± 0.04 | 0.32 ± 0.04 | 0.35 ± 0.06 | 0.39 ± 0.10 | 0.23 ± 0.06 | 0.21 ± 0.18 | 0.32 ± 0.06 | 0.12 ± 0.09 | 0.41 ± 0.20 | 0.20 ± 0.08 | 0.45 ± 0.12 | 0.27 a ± 0.14 |
| B8 | 0.59 ± 0.50 | 0.31 ± 0.04 | 0.51 ± 0.29 | 0.65 ± 0.41 | 0.66 ± 0.18 | 0.29 ± 0.24 | 0.78 ± 0.67 | 0.37 ± 0.15 | 0.36 ± 0.09 | 0.33 ± 0.06 | 0.19 ± 0.04 | 0.45 abcd ± 0.31 |
| B9 | 0.46 ± 0.32 | 0.33 ± 0.05 | 0.57 ± 0.26 | 0.85 ± 0.59 | 0.55 ± 0.42 | 0.19 ± 0.14 | 0.35 ± 0.05 | 0.04 ± 0.01 | 0.30 ± 0.03 | 0.12 ± 0.03 | 0.10 ± 0.04 | 0.34 ab ± 0.32 |
| B10 | 0.58 ± 0.10 | 0.84 ± 0.41 | 0.68 ± 0.23 | 0.77 ± 0.35 | 0.67 ± 0.08 | 0.62 ± 0.49 | 0.65 ± 0.32 | 0.49 ± 0.13 | 0.34 ± 0.11 | 0.64 ± 0.16 | 0.74 ± 0.17 | 0.64 d ± 0.26 |
| B11 | 0.62 ± 0.01 | 0.66 ± 0.26 | 0.63 ± 0.18 | 0.41 ± 0.17 | 0.71 ± 0.06 | 0.52 ± 0.23 | 0.62 ± 0.07 | 0.35 ± 0.03 | 1.00 ± 0.64 | 0.60 ± 0.05 | 0.50 ± 0.21 | 0.59 cd ± 0.28 |
| B12 | 0.47 ± 0.13 | 1.20 ± 0.03 | 1.15 ± 0.20 | 0.71 ± 0.05 | 1.30 ± 0.44 | 0.86 ± 0.44 | 0.84 ± 0.14 | 1.10 ± 0.35 | 0.75 ± 0.73 | 1.05 ± 0.07 | 0.80 ± 0.47 | 0.92 e ± 0.37 |
| B13 | 0.08 ± 0.03 | 0.35 ± 0.21 | 0.39 ± 0.15 | 0.40 ± 0.38 | 0.14 ± 0.11 | 0.25 ± 0.18 | 0.11 ± 0.07 | 0.26 ± 0.23 | 0.30 ± 0.08 | 0.16 ± 0.00 | 0.03 ± 0.02 | 0.23 a ± 0.20 |
| B14 | 0.46 ± 0.31 | 0.49 ± 0.22 | 0.89 ± 0.28 | 0.56 ± 0.09 | 1.07 ± 0.68 | 0.52 ± 0.20 | 0.48 ± 0.11 | 0.57 ± 0.09 | 0.33 ± 0.12 | 0.37 ± 0.15 | 0.16 ± 0.00 | 0.55 bcd ± 0.32 |
| Average | 0.29 a ± 0.23 | 0.44 abc ± 0.27 | 0.57 c ± 0.29 | 0.52 bc ± 0.28 | 0.53 bc ± 0.30 | 0.36 abc ± 0.33 | 0.49 abc ± 0.29 | 0.34 ab ± 0.30 | 0.51 abc ± 0.36 | 0.32 ab ± 0.30 | 0.40 abc ± 0.31 |
| L1 | L2 | L3 | L4 | L5 | L6 | L7 | L8 | L9 | L10 | L11 | Average | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| B1 | 4.86 ± 0.06 | 4.50 ± 0.09 | 4.78 ± 0.03 | 4.76 ± 0.06 | 4.77 ± 0.04 | 5.15 ± 0.85 | 4.67 ± 0.02 | 5.77 ± 0.79 | 4.37 ± 0.04 | 5.42 ± 0.07 | 4.60 ± 0.04 | 4.87 e ± 0.50 |
| B2 | 4.56 ± 0.21 | 4.30 ± 0.02 | 4.56 ± 0.08 | 4.42 ± 0.08 | 4.54 ± 0.23 | 5.10 ± 1.01 | 4.29 ± 0.03 | 5.59 ± 1.00 | 4.15 ± 0.03 | 4.98 ± 0.21 | 4.30 ± 0.06 | 4.62 cde ± 0.57 |
| B3 | 4.59 ± 0.19 | 4.06 ± 0.09 | 4.73 ± 0.17 | 4.58 ± 0.18 | 4.61 ± 0.17 | 5.78 ± 0.30 | 4.35 ± 0.07 | 5.78 ± 0.34 | 3.95 ± 0.05 | 4.71 ± 0.23 | 4.05 ± 0.06 | 4.65 cde ± 0.62 |
| B4 | 4.69 ± 0.05 | 4.15 ± 0.04 | 4.62 ± 0.22 | 4.56 ± 0.16 | 4.39 ± 0.06 | 5.65 ± 0.10 | 4.19 ± 0.06 | 6.04 ± 0.47 | 4.01 ± 0.04 | 4.75 ± 0.29 | 4.30 ± 0.06 | 4.66 cde ± 0.63 |
| B5 | 4.87 ± 0.02 | 4.63 ± 0.20 | 4.65 ± 0.10 | 4.81 ± 0.03 | 4.76 ± 0.03 | 5.26 ± 0.76 | 4.63 ± 0.03 | 5.20 ± 0.34 | 4.75 ± 0.11 | 4.80 ± 0.31 | 4.31 ± 0.04 | 4.79 de ± 0.34 |
| B6 | 4.79 ± 0.13 | 4.33 ± 0.18 | 4.55 ± 0.05 | 4.78 ± 0.13 | 4.76 ± 0.09 | 4.92 ± 0.24 | 4.43 ± 0.38 | 5.87 ± 0.88 | 3.97 ± 0.05 | 4.64 ± 0.18 | 4.18 ± 0.17 | 4.66 cde ± 0.55 |
| B7 | 5.00 ± 0.17 | 4.57 ± 0.21 | 4.69 ± 0.06 | 4.87 ± 0.04 | 4.76 ± 0.03 | 5.90 ± 0.25 | 4.69 ± 0.02 | 4.79 ± 0.48 | 4.65 ± 0.33 | 4.25 ± 0.18 | 3.97 ± 0.09 | 4.74 cde ± 0.50 |
| B8 | 5.52 ± 0.07 | 4.21 ± 0.11 | 4.44 ± 0.06 | 4.42 ± 0.27 | 4.29 ± 0.19 | 4.37 ± 0.03 | 4.20 ± 0.25 | 4.33 ± 0.07 | 4.09 ± 0.04 | 4.26 ± 0.07 | 4.22 ± 0.11 | 4.40 bcd ± 0.39 |
| B9 | 5.77 ± 0.13 | 4.33 ± 0.14 | 4.60 ± 0.07 | 4.59 ± 0.12 | 4.61 ± 0.35 | 4.98 ± 0.10 | 4.37 ± 0.08 | 4.99 ± 0.26 | 3.95 ± 0.03 | 4.88 ± 0.03 | 4.72 ± 0.18 | 4.71 cde ± 0.47 |
| B10 | 4.75 ± 0.24 | 4.10 ± 0.04 | 4.26 ± 0.05 | 4.30 ± 0.13 | 4.52 ± 0.31 | 4.46 ± 0.17 | 4.04 ± 0.08 | 4.36 ± 0.31 | 3.97 ± 0.09 | 4.60 ± 0.09 | 4.51 ± 0.07 | 4.35 abc ± 0.28 |
| B11 | 5.17 ± 0.57 | 4.11 ± 0.06 | 4.22 ± 0.02 | 4.28 ± 0.12 | 4.42 ± 0.23 | 4.62 ± 0.15 | 4.00 ± 0.04 | 4.55 ± 0.18 | 4.01 ± 0.05 | 4.44 ± 0.10 | 4.28 ± 0.15 | 4.37 abc ± 0.37 |
| B12 | 4.70 ± 0.40 | 3.90 ± 0.04 | 3.98 ± 0.03 | 4.04 ± 0.22 | 3.97 ± 0.19 | 4.08 ± 0.10 | 3.89 ± 0.08 | 4.02 ± 0.06 | 3.87 ± 0.07 | 4.15 ± 0.04 | 4.08 ± 0.18 | 4.06 ab ± 0.26 |
| B13 | 5.39 ± 0.05 | 4.48 ± 0.06 | 4.51 ± 0.03 | 4.58 ± 0.09 | 4.90 ± 0.68 | 5.47 ± 0.40 | 4.53 ± 0.03 | 5.67 ± 0.15 | 4.16 ± 0.02 | 5.36 ± 0.12 | 5.00 ± 0.23 | 4.91 e ± 0.53 |
| B14 | 5.07 ± 0.78 | 4.17 ± 0.58 | 3.93 ± 0.11 | 4.26 ± 0.91 | 3.76 ± 0.10 | 3.79 ± 0.08 | 3.73 ± 0.02 | 3.77 ± 0.04 | 3.89 ± 0.19 | 3.79 ± 0.03 | 3.81 ± 0.07 | 4.00 a ± 0.51 |
| Average | 4.97 de ± 0.44 | 4.27 ab ± 0.26 | 4.47 bc ± 0.27 | 4.52 bc ± 0.33 | 4.50 bc ± 0.38 | 4.97 de ± 0.72 | 4.29 ab ± 0.31 | 5.05 e ± 0.84 | 4.18 a ± 0.42 | 4.65 cd ± 0.46 | 4.31 ab ± 0.32 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mangieri, N.; Ambrosini, D.; Baroffio, S.; Vigentini, I.; Foschino, R.; De Noni, I. Valorisation of Bovine Sweet Whey and Sunflower Press Cake Blend through Controlled Fermentation as Platform for Innovative Food Materials. Foods 2022, 11, 1417. https://doi.org/10.3390/foods11101417
Mangieri N, Ambrosini D, Baroffio S, Vigentini I, Foschino R, De Noni I. Valorisation of Bovine Sweet Whey and Sunflower Press Cake Blend through Controlled Fermentation as Platform for Innovative Food Materials. Foods. 2022; 11(10):1417. https://doi.org/10.3390/foods11101417
Chicago/Turabian StyleMangieri, Nicola, Davide Ambrosini, Stefano Baroffio, Ileana Vigentini, Roberto Foschino, and Ivano De Noni. 2022. "Valorisation of Bovine Sweet Whey and Sunflower Press Cake Blend through Controlled Fermentation as Platform for Innovative Food Materials" Foods 11, no. 10: 1417. https://doi.org/10.3390/foods11101417